TP53在肿瘤抑制中起关键作用,50%的侵袭性肿瘤显示TP53基因突变。在这项研究中,我们描述了具有TP53缺失的结肠癌细胞(hct116p53/),它是HCT11p53+/+细胞的一个亚系。进行了RNA测序和网络分析,以确定新的耐药机制。染色体畸变通过多色荧光原位杂交和阵列比较基因组杂交(aCGH)来鉴定。许多基因在HCT116 p53细胞中过表达:rnd3/rhoe(上调235.6倍)、dclk1(上调60.2倍)、lbh(上调LBH(31.9倍)、MYB(上调MYB(28.9倍)和tacstd2(下调110.1倍)。根据以前发表的研究,NRIP1(81.5倍。可能是由于TP53的缺失,DNA修复和凋亡的紊乱,导致细胞和组织的异常,最终增加肿瘤的发生发展潜力。核因子κB、PI3K和热休克蛋白70是融合蛋白网络的中心,TH2途径是受影响的途径之一。可以推测,炎症途径、细胞周期调控和热休克反应参与了耐药表型。与HCT11p53+/+细胞相比,HCT11p53细胞染色体畸变更多,拷贝数增加和减少。综上所述,许多基因组畸变可能与未知的耐药机制有关。这可能会对未来的治疗策略产生重要影响。
简介
TP53被描述为基因组的守护者。当外来物质和致癌物质造成有害损害时,P53保持细胞完整性。DNA扭曲和损伤被p53识别,导致细胞周期停滞和DNA修复。当持续性损伤超过细胞修复能力的极限时,P53可以引发细胞凋亡。p53诱导细胞凋亡的机制包括FAS、KILLER/DR5和线粒体途径的转录激活。此外,细胞存活,如BCL2,IGFR,MCL-1,生存素和PIK3CA,被p53抑制。在翻译后修饰(如磷酸化、乙酰化和甲基化)激活后,P53参与除凋亡和生长停滞外的各种下游过程。作为一种转录因子,它可以通过调节细胞周期停滞、DNA修复和/或凋亡相关途径来维持基因组的完整性。在细胞周期调控方面,p53激活p21/waf1,p21/wa f1是G2/M特异性细胞分裂控制蛋白2激酶和细胞周期蛋白依赖性G1激酶的抑制剂,进而导致G2/M和G1检查点控制。由于突变的p53不能在G1和G2/M检查点阻止细胞,可能导致耐药性。
DNA修复和凋亡机制对维持人体细胞健康非常重要。正常情况下,过度的DNA损伤或其他扭曲的细胞是通过凋亡来消除的。如果p53启动的调节机制被破坏,就会发生异常的细胞增殖和过度的DNA损伤。P53基因突变是DNA修复和凋亡中断的主要原因之一,可能是由于异常细胞增多,更容易发生突变和染色体不稳定所致。无功能p53导致DNA损伤积累的细胞异常增殖,也会影响下一代有额外突变的细胞。最终,这将增加患癌症的风险。
超过50%的人类癌症具有P53突变,而结直肠癌是经常具有有害P53突变的癌症类型之一。大多数突变发生在DNA结合域,导致蛋白质错误折叠或破坏DNA结合能力。凋亡的丧失是放射和耐药癌细胞发展的重要原因。另外,p53突变的肿瘤通常具有加重转移和基因组不稳定的特点。突变型p53的其他致癌作用包括促进侵袭、迁移、血管生成和增殖,导致耐药性增加和有丝分裂缺陷。这些功能只是突变型p53调控癌症进展的许多方式中的一小部分。例如,p53还通过限制糖酵解和促进线粒体呼吸来影响药物代谢和细胞代谢。
在介导多药耐药表型的三磷酸腺苷结合盒转运体中,对多种药物的耐药性已被广泛研究。但多药耐药并不限于ABC转运体,其他多药耐药现象也有描述,包括p53、Bcl-2、肿瘤增殖速率等。以前已经对HCT116细胞系进行了微阵列分析,但是基因组学和转录组学在同源敲除细胞中的应用使得细胞系之间能够进行更好和更深入的比较,以识别新的耐药机制。
在这项研究中,我们使用RNA测序、阵列比较基因组杂交(aCGH)和多色荧光原位杂交(mFISH)来分析HCT116p53+/+结肠癌细胞及其p53缺失的耐药亚系HCt 116 p53/来表征基因、途径、蛋白质网络和染色体畸变与HCt 116 p53/细胞系的耐药性相关。总的来说,这项研究将概述结肠癌细胞的完整和复杂的机制和遗传变化,以及它们对p53缺失引起的耐药性的贡献。
细胞培养
HCT116 p53+/+及其耐药HCT116 p53亚系。HCT 116 p53/细胞存在明显的有丝分裂检查点缺陷,不能正常对DNA破坏剂做出反应,进入有丝分裂,然后在DNA破坏的情况下复制其基因组。在过去几年中,研究了HCT116 p53的耐药谱。与野生型细胞相比,这些敲除细胞对许多已建立的药理学抗癌药物(阿霉素、氟尿嘧啶和5’-脱氧
核糖核酸测序
基因表达通过FPKM(百万分之一千碱基转录本读取的图谱片段数)测量进行量化(Choudhri等人,2018年;;Wesolowski等人,2013年).HCT116p53+/+细胞中基因的总FPKM值除以HCT 116 p53+/+细胞中的FPKM值来计算HCT 116 p53/细胞中的去调节基因。
路径和网络分析
利用7的RNA表达的多重变化进行筛选,然后对解除调控的基因列表进行心灵手巧途径分析(IPA) (qiagen redwood city,USA,www.qiage.comn,com/心灵手巧)以鉴定hct116p53细胞中的特定网络和途径。
荧光分析系统
HCT116 p53和HCT116 p53+/+细胞根据标准程序通过细胞遗传学制备以获得中期扩散,并通过分子细胞遗传学分析。如前所述,使用人类全染色体涂料作为探针进行mFISH。
基因组检查
用QIAmp DNA微型试剂盒从HCT 116 p53/-和HCT116 p53+/+细胞中提取全基因组DNA。ACGH沿用之前的报告。
蛋白质印迹
评估HCT116 p53和HCT116 p53+/+细胞中选定基因(即ANGPT2和过氧化氢酶)的蛋白表达水平,以验证通过RNA测序分析发现的去调控。使用蛋白质提取缓冲液(M-PER)从细胞中提取总蛋白质,哺乳动物蛋白质提取试剂和1%Halt混合蛋白酶抑制剂的混合物。将相当于30克的样品装入10%的SDS-PAGE中进行分离,然后转移到Ruti-聚偏氟乙烯膜上。膜用5%牛血清白蛋白封闭1小时,在4℃(1∶1000)下用选定的抗血管生成素样蛋白2、过氧化氢酶和β-肌动蛋白的初级抗体检测。24小时后,与辣根过氧化物酶二抗(1:2000)孵育1小时,Luminata检测经典的蛋白质辣根过氧化物酶底物。ImageJ软件用于图像分析。
结果
HCT116 p53细胞差异基因表达谱、下游途径及网络分析
与HCT116 p53+/+细胞相比,HCT116 p5细胞中各基因表达的RNA-seq衍生FPKM值的比值被认为是基因表达的倍数变化。为了进一步分析,考虑了阈值为7的差异基因表达,获得了300个差异表达基因。表1列出了HCT11p53细胞中前10个上调和下调的基因。RND3(+235.6)、MCPH1(+85.7)和MYB(+28.9)是上调最多的基因,而DPEP1(431.9)、ICAM 1 (166.5)和NPM 2(85.4)是下调最多的基因。
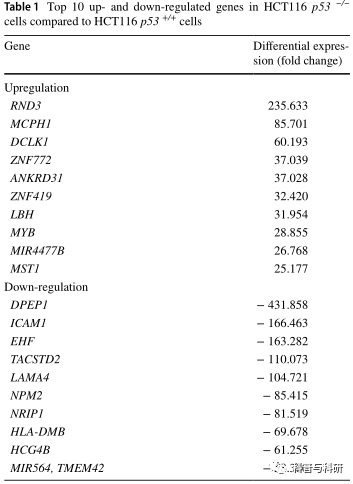
表1与HCT116 p53 +/+细胞相比,HCT116 p53 +/细胞中前10位上调和下调基因
在网络1中,组蛋白H4、细胞周期蛋白a和核因子κB的节点最多,CD3和Hsp70的节点最多。网络1中的“癌症”、“组织损伤和异常”以及网络2中的“细胞组装和组织”和“分子转运”是受影响的生物功能。Erk1/2是网络3中节点数量最多的。“损伤和异常”和“碳水化合物代谢”是网络3中受影响的生物功能(图1)。


图1 HCT 116 p53/-细胞中受影响的蛋白质网络与HCT116p53 +/+细胞的比较。标记绿色的基因下调,标记红色的基因上调。
一些已知参与耐药性的基因被解除调控,这意味着HCT116 p53细胞显示多因子耐药性表型。一个DNA修复基因、一个氧化应激基因和一个转录因子基因属于解除调控的耐药基因,这意味着这些基因可能对HCT116 p53的MDR表型有重要影响。这些基因如表2所示,所有参与抗性机制的去调控基因的完整列表如补充表2所示。

表2与野生型HCT116 p53 +/+细胞相比,HCT 116 p53+/+细胞中参与经典耐药机制的去调节基因(阈值:表达改变7倍)
合并前三个网络,并进一步分析合并后的网络。(图2)显示核因子κB与PI3K和热休克蛋白70一起位于融合网络的中心。

图2网络1、2和3的组合蛋白质网络
“癌症”、“身体损伤和异常”和“细胞间信号和相互作用”是HCT116p53细胞中受影响最大的生物学功能(图3)。位于前10位生物功能表中的基因如补充表3所示。

图3与HCT116 p53 +/+细胞相比,HCT116 p53 +/细胞中受影响的生物学功能(前10)。橙色线表示统计显著性阈值(p = 0.05)
Th1通路(p值:0.000437)和IL4信号通路(p值:0.012589)是HCT116 p53细胞中最重要的信号通路之一,这意味着可能的免疫反应通路对耐药性有影响(图4)。

图4。与HCT116 p53 +/+细胞相比,HCT116 p53 +/细胞中受影响的信号通路(前10)。橙色线描绘了统计显著性阈值(p = 0.05),橙色图描绘了每个途径中去调节基因的比例
在116个p53靶基因中(Fischer 2017),33个下调,19个上调(倍数变化高于阈值1.5),如表3所示。


表3与HCT11p53 +/+细胞相比,HCT 116 p53/-细胞中p53靶基因的去调控
在蛋白质水平上,验证了所选基因的血管生成素样蛋白2和过氧化氢酶。如图5所示,hct116p53细胞中ANGPT2上调(+8.7倍),过氧化氢酶下调(1.9倍),与RNA测序输出有关,RNA表达数据在蛋白质水平得到验证。
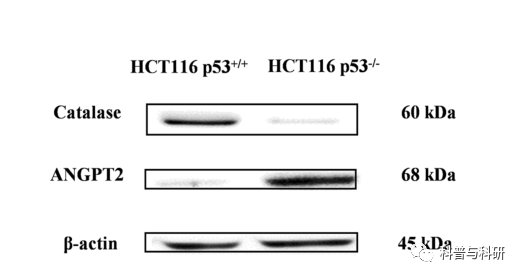
图5通过蛋白质印迹法测定HCT116 p53和HCT116p53 +/+细胞中血管生成素样蛋白2和过氧化氢酶的表达
HCT11p53 +/+细胞核型为45


图6:HCT 116 p53+/+(a)和hct116p53/-(b)细胞的mFISH分析
比较基因组杂交
HCT116 p53 +/+:染色体扩增和缺失很好地反映在RNA Seq分析中观察到的基因表达的去调控中。LVRN下调3.3倍,AP3S1上调1.7倍。相应的染色体基因座(5q23.1)被扩增。FLJ42393下调1.7倍。在相应的染色体位点(3q 27.3–q28)有一个缺失。结果总结在表4中。HCT116 p53/:与HCT116 p53 +/+细胞相比,在HCT 11p53/+细胞中观察到更多的扩增和缺失。这意味着p53缺失导致额外的染色体畸变、扩增和缺失积累。NXPH2表达上调1.7倍,并在相应的染色体位点扩增。OR5K2表达上调1.7倍,并在相应的染色体位点扩增。PARK2下调8.1倍,在相应的染色体位点有一个缺失。aCGH数据和RNA-Seq结果之间的相关性清楚地显示了基因在相应染色体位点的差异表达,如表4所示。表5描述了aCGH的总体结果。
表4染色体畸变和相应的去调控基因。aCGH和核糖核酸测序图的比较


表5:加拿大全球健康调查总体结果

在本研究中,结肠癌细胞系HCT116 p53+/+和具有TP53缺失的耐药HCT116 p53亚系被用作识别新的耐药基因的模型。基因表达谱、信号通路、生物学功能和染色体异常通过核糖核酸测序、荧光原位杂交和aCGH鉴定。一些已知与耐药性有关的基因被解除调控,支持HCT116 p53细胞的多因子耐药性表型。这些基因在各种耐药性簇中被鉴定,包括凋亡、DNA修复、铁下垂、谷胱甘肽相关、热休克和氧化应激。表2中列出的转录因子可能对MDR表型有重要影响。
上调最多的基因RND3/RhoE(+235.6倍)以前与肿瘤侵袭和转移有关,并被报道为胃癌和结直肠癌病例的肿瘤耐药性、复发和预后的潜在标志。CARD11的突变与布洛芬有关。CARD11(7.4倍)出现在前五个具有生物功能的基因中,表明凋亡抑制下调的CARD 11可能在HCT116p53细胞的耐药表型中起重要作用。据报道,CARD11导致了布洛芬在癌症中的耐药性,这意味着CARD11可能在HCT 116 P 53/细胞的耐药性表型中起作用。据报道,DCLK1(+60.2倍)与非小细胞肺癌细胞的顺铂耐药性有关,靶向DCLK1的miR539导致对顺铂的敏感性增加。DCLK1还与结直肠癌、胰腺癌和肾癌的耐药性有关。据报道,LBH(+31.9倍)是肝细胞癌的潜在标志,因为其过度表达与不良预后有关。Myb(+敲除细胞表达28.9倍)是一种致癌转录因子,对白血病细胞的转化起促进作用。Myb与结肠癌细胞对顺铂的耐药性有关。它还参与一些实体瘤的发展和进展,包括黑色素瘤中TACSD 2的缺失,TACSD 2通过抑制化疗药物诱导的凋亡而促进鳞状细胞癌的进展和耐药性,这意味着TACSD 2可以作为鳞状细胞癌病理分级的标志(图1,表2,与野生型HCT116 p53+/+细胞相比,在HCT116 p5细胞中,参与经典耐药机制(阈值:7倍变化表达)的去调节基因差异表达(倍变化)。DNA修复XRCC211.200 BRIP1 7.711氧化应激NCF 215.131 MB 8.093转录因子myb28.855 NFATC47.619 .在HCT 116 p53/细胞中下调110.1倍,表明TACSTD2的下调可能是导致HCT 116 p53/细胞侵袭性生长和多药耐药的机制。NRIP1下调后,食管鳞状细胞癌细胞的迁移和侵袭能力增强。在这项研究中,观察到NRIP1在HCT116p53细胞中下调81.5倍,这意味着NRIP1的下调可能在多药耐药表型中起作用。人类白细胞抗原-DMB (69.7倍)属于主要组织相容性复合体ⅱ类基因。在晚期浆液性卵巢癌中,人类白细胞抗原-DMB的高表达通过增加CD8淋巴细胞的数量与较高的生存率有关。HLADMB的下调可能通过影响肿瘤侵袭性与HCT116 p53细胞的MDR表型有关。
MYB转录因子的过度表达(+敲除细胞表达的28.8倍)与预后不良相关,并且经常在结直肠癌中观察到。另一项研究指出,MYB在肿瘤细胞中的表达由于其致瘤性而调节宿主免疫反应,这可能影响对结直肠癌患者的免疫治疗的使用。MYC(+5.1倍)是另一种在肿瘤发生中起关键作用的转录因子。它调节细胞周期相关基因的表达,并已在包括结肠癌在内的各种癌症类型中观察到。
在本实验中,在hct116p53细胞中鉴定出IL4信号通路,这意味着免疫反应通路可能影响耐药性。一项研究表明,白细胞介素-4可以增强BCR信号,降低BCR激酶抑制剂(如布洛芬)在CLL细胞中的效力。另一项研究报道,白细胞介素-1受体相关激酶1和4(IRAK1/4)复合物激活先天免疫途径有助于FLT3突变AML细胞的适应性抵抗。这一结果支持了免疫反应途径和耐药性之间的关系。
RNA-seq结果通过ANGPT2和过氧化氢酶的westernblotting验证。与HCT 116 p5/+细胞相比,angpt2mrna在HCT116 p5/-细胞中上调,而CAT mRNA在HCT 116 p5/-细胞中下调。这一点通过蛋白质表达得到了证实。CAT在肿瘤中经常下调。例如,乳腺癌的特征是过氧化氢酶的下调和超氧化物歧化酶的过度表达。另一方面,ANGPT2的上调与结肠癌肝转移有关。
根据网络分析,“癌症”、“机体损伤与异常”、“细胞组装与组织”、“分子转运”、“机体损伤与异常”、“碳水化合物代谢”是主要影响因素。由于TP53缺失,破坏DNA修复和凋亡机制,可能导致细胞和组织结构异常,最终增加肿瘤的发生发展潜力。在网络1中,编码组蛋白H4、细胞周期蛋白A和核因子κB的基因具有最高的节点数。CD3和HSP70在网络2中心节点最多,说明细胞周期调控影响炎症,热休克反应影响耐药性。重要的是,网络2中分子转运蛋白的出现,强调了p53与细胞转运蛋白可能的相互作用,从而促进癌细胞的MDR。ABB1/MDR 1基因是ABC转运蛋白的成员,在转录中依赖p53,野生型p53通过与下游启动子的序列特异性结合对ABB1/MDR 1基因表达产生负面影响。相反,突变型p53在不同细胞系中激活ABCB1/MDR1启动子。ERK1/2基因在网络3中节点数最多,说明ERK调节的细胞增殖途径有助于耐药。
核因子κB与PI3K和热休克蛋白70一起位于融合网络的中心,这意味着炎症途径、细胞周期和热休克反应在多药耐药表型中的作用。Th1、Th2通路和CD28信号通路是HCT11p5细胞中受影响最大的信号通路之一,支持炎症通路在MDR表型中起重要作用的假说。
结论
总的来说,通过RNA测序、mFISH和aCGH分析结肠癌细胞系HCT116 p53和HCT116p53 +/+的基因表达谱,通过比较鉴定差异表达基因、受影响的蛋白质网络、途径、生物学功能和染色体畸变。已经发现了多种基因、途径和网络,它们可能与结肠癌的耐药性和侵袭行为有关。这项研究清楚地表明,TP53敲除细胞的耐药性是由许多因素决定的,而不是由单个因素决定的。显然,多因素耐药性使新治疗策略的开发变得复杂。然而,这项研究可能代表了设计更具体和更有希望的抗癌策略以绕过耐药性的起点。
投稿人:多多
审计:黛西
排版:安静